
La célula eucariota

Diagrama de una célula animal, a la izquierda (1. Nucléolo, 2. Núcleo, 3. Ribosoma, 4. Vesícula, 5. Retículo endoplasmático rugoso, 6. Aparato de Golgi, 7. Citoesqueleto (microtúbulos), 8. Retículo endoplasmático liso, 9. Mitocondria, 10. Vacuola, 11. Citoplasma, 12. Lisosoma. 13. Centríolos.); y de una célula vegetal, a la derecha.
Compartimentos
Las células son entes dinámicos, con un metabolismo celular interno de gran actividad cuya estructura es un flujo entre rutas anastomosadas. Un fenómeno observado en todos los tipos celulares es la compartimentalización, que consiste en una heterogeneidad que da lugar a entornos más o menos definidos (rodeados o no mediante membranas biológicas) en las cuales existe un microentorno que aglutina a los elementos implicados en una ruta biológica.[37] Esta compartimentalización alcanza su máximo exponente en las células eucariotas, las cuales están formadas por diferentes estructuras y orgánulos que desarrollan funciones específicas, lo que supone un método de especialización espacial y temporal.[1] No obstante, células más sencillas, como los procariotas, ya poseen especializaciones semejantes.[38]
Membrana plasmática y superficie celular
Artículo principal: Membrana plasmática
La composición de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentre, pero posee elementos comunes. Está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y por glúcidos unidos covalentemente a lípidos o proteínas. Generalmente, las moléculas más numerosas son las de lípidos; sin embargo, la proteínas, debido a su mayor masa molecular, representan aproximadamente el 50% de la masa de la membrana.[37]
Un modelo que explica el funcionamiento de la membrana plasmática es el modelo del mosaico fluido, de J. S. Singer y Garth Nicolson (1972), que desarrolla un concepto de unidad termodinámica basada en las interacciones hidrófobas entre moléculas y otro tipo de enlaces no covalentes.[39]

Esquema de una membrana celular. Se observa la bicapa de fosfolípidos, las proteínas y otras moléculas asociadas que permiten las funciones inherentes a este orgánulo.
Dicha estructura de membrana sustenta un complejo mecanismo de transporte, que posibilita un fluido intercambio de masa y energía entre el entorno intracelular y el externo.[37] Además, la posibilidad de transporte e interacción entre moléculas de células aledañas o de una célula con su entorno faculta a éstas poder comunicarse químicamente, esto es, permite la señalización celular. Neurotransmisores, hormonas, mediadores químicos locales afectan a células concretas modificando el patrón de expresión génica mediante mecanismos de transducción de señal.[40]
Sobre la bicapa lipídica, independientemente de la presencia o no de una pared celular, existe una matriz que puede variar, de poco conspicua, como en los epitelios, a muy extensa, como en el tejido conjuntivo. Dicha matriz, denominada glucocalix (glicocáliz), rica en líquido tisular, glucoproteínas, proteoglicanos y fibras, también interviene en la generación de estructuras y funciones emergentes, derivadas de las interacciones célula-célula.[11]
Estructura y expresión génica
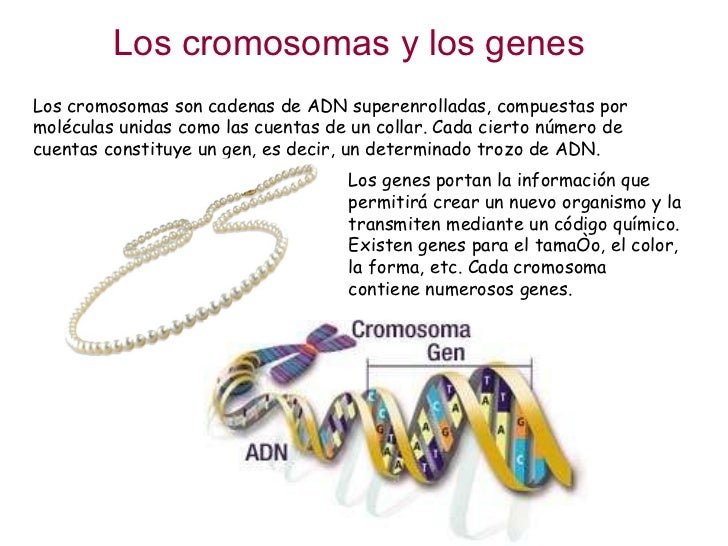
Dicho material genético se encuentra inmerso en una actividad continua de regulación de la expresión génica; las ARN polimerasas transcriben ARN mensajero continuamente, que, exportado al citosol, es traducido a proteína, de acuerdo a las necesidades fisiológicas. Asimismo, dependiendo del momento del ciclo celular, dicho ADN puede entrar en replicación, como paso previo a la mitosis.[33] No obstante, las células eucarióticas poseen material genético extranuclear: concretamente, en mitocondrias y plastos, si los hubiere; estos orgánulos conservan una independencia genética parcial del genoma nuclear.[42] [43]
Síntesis y degradación de macromoléculas
Dentro del citosol, esto es, la matriz acuosa que alberga a los orgánulos y demás estructuras celulares, se encuentran inmersos multitud de tipos de maquinaria de metabolismo celular: orgánulos, inclusiones, elementos del citoesqueleto, enzimas... De hecho, estas últimas corresponden al 20% de las enzimas totales de la célula.[11]
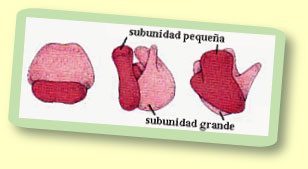
Estructura de los ribosomas; 1,: subunidad mayor, 2: subunidad menor.

Imagen de un núcleo, el retículo endoplasmático y el aparato de Golgi; 1, Núcleo. 2, Poro nuclear.3, Retículo endoplasmático rugoso (REr).4, Retículo endoplasmático liso (REl). 5, Ribosoma en el RE rugoso. 6, Proteínas siendo transportadas.7, Vesícula (transporte). 8, Aparato de Golgi. 9, Lado cis del aparato de Golgi.10, Lado trans del aparato de Golgi.11, Cisternas del aparato de Golgi.
- Ribosoma: Los ribosomas, visibles al microscopio electrónico como partículas esféricas,[44] son complejos supramoleculares encargados de ensamblar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero. Elaborados en el núcleo, desempeñan su función de síntesis de proteínas en el citoplasma. Están formados por ARN ribosómico y por diversos tipos de proteínas. Estructuralmente, tienen dos subunidades. En las células, estos orgánulos aparecen en diferentes estados de disociación. Cuando están completos, pueden estar aislados o formando grupos (polisomas). También pueden aparecer asociados al retículo endoplasmático rugoso o a la envoltura nuclear.[33]
- Retículo endoplasmático: El retículo endoplasmático es orgánulo vesicular interconectado que forma cisternas, tubos aplanados y sáculos comunicados entre sí. Intervienen en funciones relacionadas con la síntesis proteica, glicosilación de proteínas, metabolismo de lípidos y algunos esteroides, detoxificación, así como el tráfico de vesículas. En células especializadas, como las miofibrillas o células musculares, se diferencia en el retículo sarcoplásmico, orgánulo decisivo para que se produzca la contracción muscular.[13]
Aparato de Golgi: El aparato de Golgi es un orgánulo formado por apilamientos de sáculos denominados dictiosomas, si bien, como ente dinámico, éstos pueden interpretarse como estructuras puntuales fruto de la coalescencia de vesículas.[45] [46] Recibe las vesículas del retículo endoplasmático rugoso que han de seguir siendo procesadas. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos y la síntesis de polisacáridos de la matriz extracelular. Posee tres compartimientos; uno proximal al retículo endoplasmático, denominado «compartimento cis», donde se produce la fosforilación de las manosas de las enzimas que han de dirigirse al lisosoma; el «compartimento intermedio», con abundantes manosidasas y N-acetil-glucosamina transferasas; y el «compartimento o red trans», el más distal, donde se transfieren residuos de galactosa y ácido siálico, y del que emergen las vesículas con los diversos destinos celulares.[11]
- Lisosoma: Los lisosomas son orgánulos que albergan multitud de enzimas hidrolíticas. De morfología muy variable, no se ha demostrado su existencia en células vegetales.[11] Una característica que agrupa a todos los lisosomas es la posesión de hidrolasas ácidas: proteasas, nucleasas, glucosidasas, lisozima, arilsulfatasas, lipasas, fosfolipasas y fosfatasas. Procede de la fusión de vesículas procedentes del aparato de Golgi, que, a su vez, se fusionan en un tipo de orgánulo denominado endosoma temprano, el cual, al acidificarse y ganar en enzimas hidrolíticos, pasa a convertirse en el lisosoma funcional. Sus funciones abarcan desde la degradación de macromoléculas endógenas o procedentes de la fagocitosis a la intervención en procesos de apoptosis.[47]
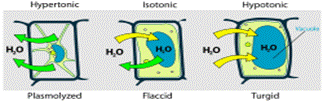
La vacuola regula el estado de turgencia de la célula vegetal.
- Vacuola vegetal: Las vacuolas vegetales, numerosas y pequeñas en células meristemáticas y escasas y grandes en células diferenciadas, son orgánulos exclusivos de los representantes del mundo vegetal. Inmersas en el citosol, están delimitadas por el tonoplasto, una membrana lipídica. Sus funciones son: facilitar el intercambio con el medio externo, mantener la turgencia celular, la digestión celular y la acumulación de sustancias de reserva y subproductos del metabolismo.[36]
- Inclusión citoplasmática: Las inclusiones son acúmulos nunca delimitados por membrana de sustancias de diversa índole, tanto en células vegetales como animales. Típicamente se trata de sustancias de reserva que se conservan como acervo metabólico: almidón, glucógeno, triglicéridos, proteínas... aunque también existen de pigmentos.[11]
Conversión energética
El metabolismo celular está basado en la transformación de unas sustancias químicas, denominadas metabolitos, en otras; dichas reacciones químicas transcurren catalizadas mediante enzimas. Si bien buena parte del metabolismo sucede en el citosol, como la glucólisis, existen procesos específicos de orgánulos.[40]

Modelo de una mitocondria: 1, membrana interna; 2, membrana externa; 3, cresta mitocondrial; 4, matriz mitocondrial.
- Mitocondria: Las mitocondrias son orgánulos de aspecto, número y tamaño variable que intervienen en el ciclo de Krebs, fosforilación oxidativa y en la cadena de transporte de electrones de la respiración. Presentan una doble membrana, externa e interna, que dejan entre ellas un espacio perimitocondrial; la membrana interna, plegada en crestas hacia el interior de la matriz mitocondrial, posee una gran superficie. En su interior posee generalmente una sola molécula de ADN, el genoma mitocondrial, típicamente circular, así como ribosomas más semejantes a los bacterianos que a los eucariotas.[11] Según la teoría endosimbiótica, se asume que la primera protomitocondria era un tipo de proteobacteria.[48]
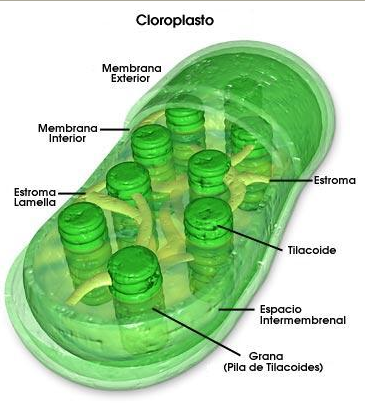
Estructura de un cloroplasto.
- Cloroplasto: Los cloroplastos son los orgánulos celulares que en los organismos eucariotas fotosintéticos se ocupan de la fotosíntesis. Están limitados por una envoltura formada por dos membranas concéntricas y contienen vesículas, los tilacoides, donde se encuentran organizados los pigmentos y demás moléculas implicadas en la conversión de la energía luminosa en energía química. Además de esta función, los plastidios intervienen en el metabolismo intermedio, produciendo energía y poder reductor, sintetizando bases púricas y pirimidínicas, algunos aminoácidos y todos los ácidos grasos. Además, en su interior es común la acumulación de sustancias de reserva, como el almidón.[11] Se considera que poseen analogía con las cianobacterias.[49]
- Peroxisoma: Los peroxisomas son orgánulos muy comunes en forma de vesículas que contienen abundantes enzimas de tipo oxidasa y catalasa; de tan abundantes, es común que cristalicen en su interior. Estas enzimas cumplen funciones de detoxificación celular. Otras funciones de los peroxisomas son: las oxidaciones flavínicas generales, el catabolismo de las purinas, la beta-oxidación de los ácidos grasos, el ciclo del glioxilato, el metabolismo del ácido glicólico y la detoxificación en general.[11] Se forman de vesículas procedentes del retículo endoplasmático.[50]
Citoesqueleto
Las células poseen un andamiaje que permite el mantenimiento de su forma y estructura, pero más aún, éste es un sistema dinámico que interactúa con el resto de componentes celulares generando un alto grado de orden interno. Dicho andamiaje está formado por una serie de proteínas que se agrupan dando lugar a estructuras filamentosas que, mediante otras proteínas, interactúan entre ellas dando lugar a una especie de retículo. El mencionado andamiaje recibe el nombre de citoesqueleto, y sus elementos mayoritarios son: los microtúbulos, los microfilamentos y los filamentos intermedios.[1] [b]
- Microfilamentos: Los microfilamentos o filamentos de actina están formados por una proteína globular, la actina, que puede polimerizar dando lugar a estructuras filiformes. Dicha actina se expresa en todas las células del cuerpo y especialmente en las musculares ya que está implicada en la contracción muscular, por interacción con la miosina. Además, posee lugares de unión a ATP, lo que dota a sus filamentos de polaridad.[51] Puede encontrarse en forma libre o polimerizarse en microfilamentos, que son esenciales para funciones celulares tan importantes como la movilidad y la contracción de la célula durante la división celular.[45]
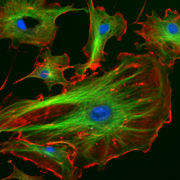
Citoesqueleto eucariota: microfilamentos en rojo, microtúbulos en verde y núcleo en azul.
- Microtúbulos: Los microtúbulos son estructuras tubulares de 25 nm de diámetro exterior y unos 12 nm de diámetro interior, con longitudes que varían entre unos pocos nanómetros a micrómetros, que se originan en los centros organizadores de microtúbulos y que se extienden a lo largo de todo el citoplasma. Se hallan en las células eucariotas y están formadas por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina. Las tubulinas poseen capacidad de unir GTP.[1] [45] Los microtúbulos intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis) y que, junto con los microfilamentos y los filamentos intermedios, forman el citoesqueleto. Además, constituyen la estructura interna de los cilios y los flagelos.[1] [45]
- Filamentos intermedios: Los filamentos intermedios son componentes del citoesqueleto. Formados por agrupaciones de proteínas fibrosas, su nombre deriva de su diámetro, de 10 nm, menor que el de los microtúbulos, de 24 nm, pero mayor que el de los microfilamentos, de 7 nm. Son ubicuos en las células animales, y no existen en plantas ni hongos. Forman un grupo heterogéneo, clasificado en cinco familias: las queratinas, en células epiteliales; los neurofilamentos, en neuronas; los gliofilamentos, en células gliales; la desmina, en músculo liso y estriado; y la vimentina, en células derivadas del mesénquima.[11]

Micrografía al microscopio electrónico de barrido mostrando la superficie de células ciliadas del epitelio de los bronquiolos.
- Centríolos: Los centríolos son una pareja de estructuras que forman parte del citoesqueleto de células animales. Semejantes a cilindros huecos, están rodeados de un material proteico denso llamado material pericentriolar; todos ellos forman el centrosoma o centro organizador de microtúbulos que permiten la polimerización de microtúbulos de dímeros de tubulina que forman parte del citoesqueleto. Los centríolos se posicionan perpendicularmente entre sí. Sus funciones son participar en la mitosis, durante la cual generan el huso acromático, y en la citocinesis,[52] así como, se postula, intervenir en la nucleación de microtúbulos.[53] [54]
- Cilios y flagelos: Se trata de especializaciones de la superficie celular con motilidad; con una estructura basada en agrupaciones de microtúbulos, ambos se diferencian en la mayor longitud y menor número de los flagelos, y en la mayor variabilidad de la estructura molecular de estos últimos.[11]

{kind=link}
{kind=link}
Es un orgánulo propio de la célula eucariota que participa en la síntesis y transporte de proteínas. El retículo endoplasmático rugoso está formado por una serie de canales que se encuentran distribuidos por todo el citoplasma de la célula. Son sacos aplanados por los que circulan todas las proteínas de la célula antes de ir al Aparato de Golgi. Existe una conexión física entre el retículo endoplasmático rugoso y el retículo endoplasmático liso.
Retículo endoplasmático liso
El retículo endoplasmático liso es un organoide celular formado por cisternas, tubos aplanados y sáculos membranosos que forman un sistema de tuberías que participa en el transporte celular y en la síntesis de triglicéridos, fosfolípidos y esteroides. También dispone de enzimas destoxificantes, que metabolizan el alcohol y otras sustancias químicas. Tiene como función la síntesis de lípidos.